*** minoken ***
![]() 中央大学基幹理工学部生命科学科
中央大学基幹理工学部生命科学科
中央大学大学院理工学研究科生命科学専攻
細胞機能学研究室(箕浦研究室)
![]()
Home > 研究材料:クラミドモナス
クラミドモナス Chlamydomonas reinhardtii(和名:コナミドリムシ)

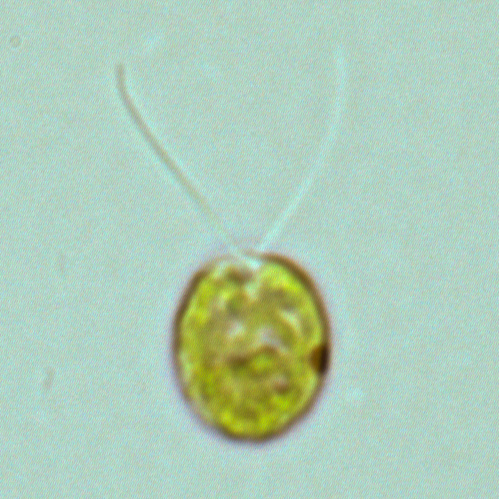 クラミドモナスは、緑藻(Chlorophyta)に属する単細胞性の生物であ
クラミドモナスは、緑藻(Chlorophyta)に属する単細胞性の生物であ
る。19世紀後半ドイツの動物学者Ehrenbergによって、ギリシア語でマン
トを意味する chlamys にちなんで名付けられた。その後、研究用株とし
て米国マサチューセッツ州のジャガイモ畑から採取された
Chlamydomonas reinhardtii 137C株が、今日に至るまで、
世界中の研究室で用いられている。
クラミドモナス
体長はおよそ10 μm
以下に挙げる多くの利点をもつことから、クラミドモナスはモデル生物のひとつとして
様々な研究に用いられている。
Chlamysをまとう旅人
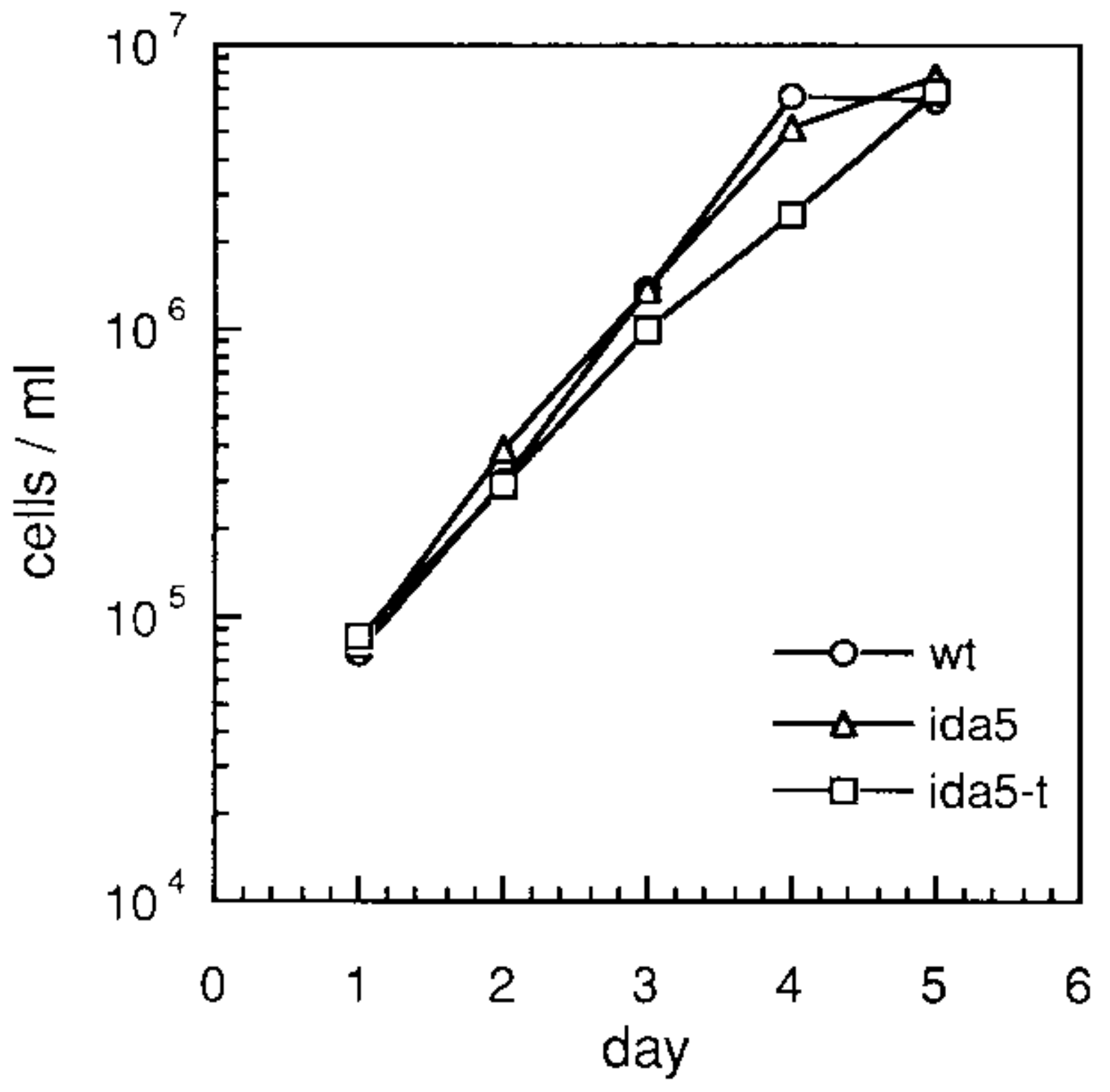
・培養が容易で、ライフサイクルが短い(当研究室では通常、25℃
前後、12時間明/12時間暗のサイクルで培養している。この場合、
細胞数は1日あたりおよそ4倍に増える)。
クラミドモナスの増殖速度
(Kato-Minoura et al., 1997)
・比較的単純な組成の培地で、液体培地でも寒天培地でも培養が可能。
・通常は核相が単相(n)なので、遺伝子に生じた変異が直ちに表現型として現れることから、
突然変異株を得やすい。
・接合型(+)と接合型(ー)の株の間で、有性生殖(接合)を行う。接合は、培地の窒素源
を除く、光条件をコントロールするなどの操作により実験室で容易に誘導できる。これによ
り、二重変異株の作成や各種の遺伝学的解析が可能。
クラミドモナスの生活環
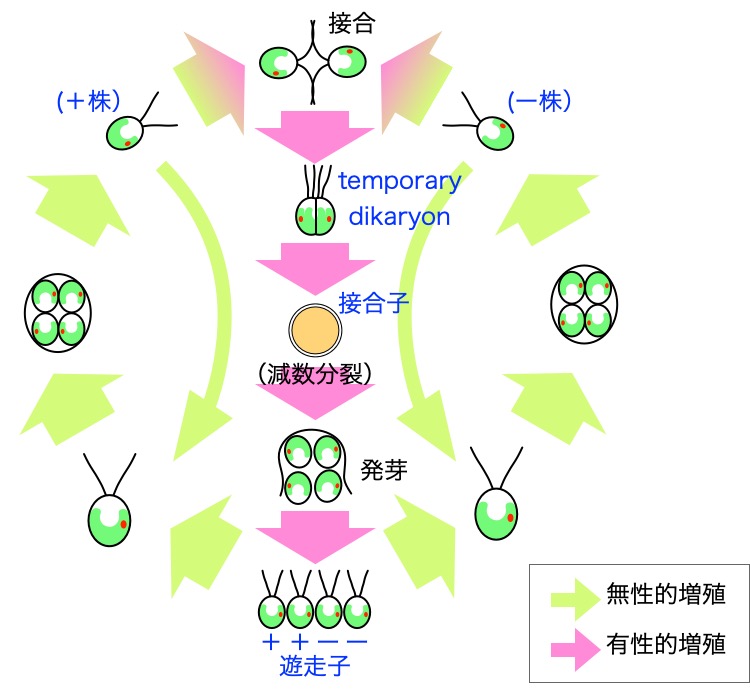
窒素源を豊富に含む培地では、クラミドモナスは無性
的に増殖する。明暗周期下で培養すると、明期には遊泳
しながら光合成を行う。暗期には二分裂を行う。
窒素源が枯渇すると、栄養細胞(遊走子、vegetative
cells)が配偶子(gametes)に分化する。そして、接
合型(+)の株と接合型(ー)の株が接合し、接合子
(zygotes)となる。
クラミドモナスの生活環



培養の様子:
液体培地での通気培養、寒天培地での培養、低温(15℃)・弱光での長期保存が可能
クラミドモナスの細胞内構造
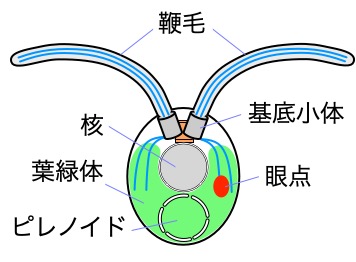
私たちの研究グループでは、チューブリンやアクチンの他、ダイニンなど様々な鞭毛構成タンパク質の突然変異株を用いて、それらの構造や機能を理解するための研究を行なっています。
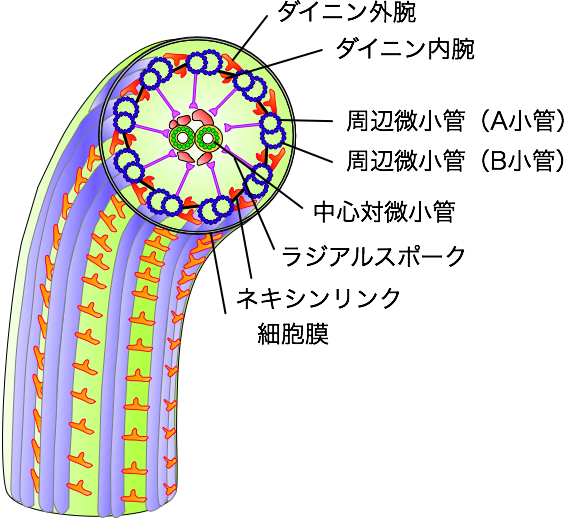
鞭毛の内部構造

野生株の鞭毛軸糸断面
(電子顕微鏡観察・負染色)
(Kato et al., 1993)
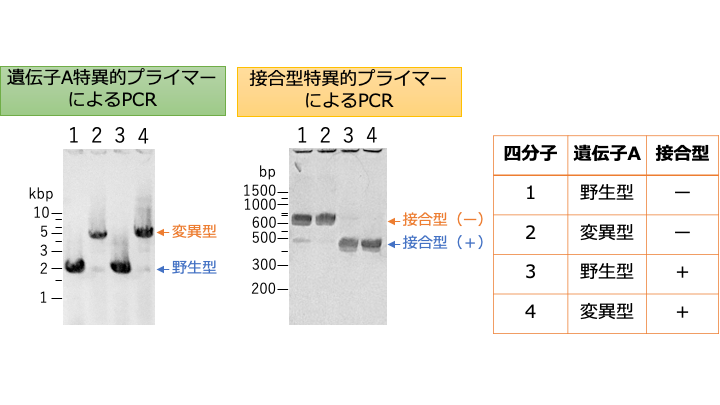
四分子解析とPCRによる遺伝子型の判定
任意の2株を掛け合わせて接合子を得たのち、四分子解析により、求める遺伝子型をもつ次世代株を得る。一例として、遺伝子Aの変異株と野生株との掛け合わせ(戻し交配)により、接合型の異なる遺伝子Aの変異株(2世代目)を得る方法を下に示す。
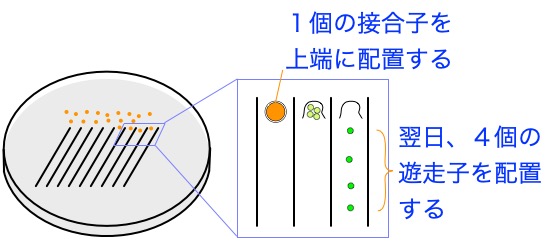
・寒天プレートに等間隔で線を引き、極細のガラス針を用い
て、線の上端に接合子を1つずつ並べる。
・翌日(およそ20時間後)、発芽した4個の遊走子
(tetrad)を十分な間隔をあけて配置する。
クラミドモナスの全ゲノム情報
クラミドモナスの全ゲノム配列は、2007年に解読・発表された。現在、JGI(Joint Genome Institute)において公開されている。
Phytozomeクラミドモナス全ゲノムデータベース(v5.6)
・系統:C. reinhardtii CC503 mt(+)
・ゲノムサイズ:111.1 Mb
・染色体 17 本+37 scaffolds
・ゲノムの99.5%をカバー
・うち、タンパク質をコードする遺伝子数:〜17000
Merchant, S. S., et al. (2007) The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science, 318(5848), 245–250.
アクチン遺伝子(Cre13.g603700)の例
・ID: Cre13.g603700
・第13染色体上(順方向に配置)
・遺伝子座:IDA5
・アクチン、内腕ダイニン(a, b, c, d, e, fとその他のマイナー種)のサブユニット
・(ー)型配偶子により発現誘導される
CLiP (Chlamydomonas Library Project)
CLiPは、クラミドモナスCC-4533 (cw15, mt-) 株に、マーカー遺伝子カセット(CIB1カセット)をランダムに挿入することによって得られた突然変異株コレクションである。2016年現在で37000株が登録されており、クラミドモナスのほとんどの遺伝子をカバーしている。
CIB1カセットの構造
選択マーカーとなるパロモマイシン耐性遺伝子(AphVIII)と、2箇所の内部バーコード(IB)をもつ。
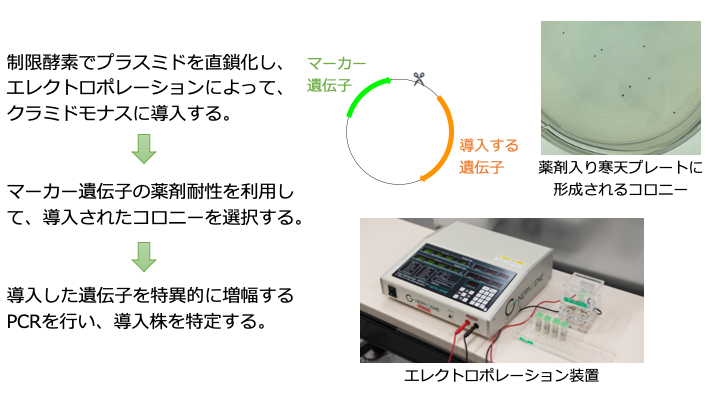
遺伝子導入