真 核生物のベン毛と繊毛
上村慎治
2008.1.15
らせん状繊維が基部にあるモーターで回転するのがバ クテリアのベン毛であるのに対して、真核生物のベン毛・繊毛は、それ自身で能動的なくねりを運動する。骨格筋では、大脳や脊髄などの神経系があってはじめ て整然とした動きができるのに対して、ベン毛・繊毛は、自前の制御機構をすべて持っている点がユニークである。一旦、何らかの信号刺激によって運動がス タートすると、エネルギーの供給がある限り、自律的なくねり運動を続けることができる。多少の荷重負荷の変化や障害物にも対処できる。太さ0.2ミクロン、長さが数〜数十ミクロンの小さな運動器官であるが、自律マシンとしては、なかなかのすぐれ物であると思う。 最近のこの研究分野の動向や進化の歴史に関する見解などを交えて、この数年の私の研究室の仕事を紹介する。
ベン毛・繊毛の1つの大きな特長は、その進化の歴史である。おそらく真核生物の持つ最も古い運動器官の1つではないかと考えられている(1)。15〜18億年
前、私たちの祖先が単細胞であった頃、ベン毛や繊毛はすでに現在の完成形に近い形で存在していたと考えられて
いる(図1、図 S-1注)。
その時点で2つに別れたグループ、ユニコンタ(1本のベン毛を持つ動物・アメーバなどの祖先)とバイコ
ンタ(2本のベン毛を持つ高等植物・繊毛虫などの祖先)の両者に共通した9+2構造(図2、図S-2)があり、同じモータータンパク質(ダイニン)による同じ様式の運動が見られるからである。その後、高等植物やアメーバなどでは、何らかの理
由でベン毛・繊毛が必要なくなり、ダイニン分子もゲノム情報の中から完全に消失した(2)。逆に、我々ヒトは、その
祖先型の姿を失っておらず、精子の運動や気管
や輸卵管での物質輸送のしくみとして役だっている。最近の研究から、私たちの体を作るほとんどすべての細胞が、常に、あるいは、何らかの条件下で、繊毛を
1本持った祖先型ユニコンタのような細胞形態となり(この場合、プライマリー繊毛と呼ぶ)、これが重要な機能発現や分化の引き金となっているのではないか
と考えられるようになった。ベン毛・繊毛の機能に関係したタンパク質の欠失で、重篤な症状となる遺伝病が多数あることがわかって来たからである(3,4,5)。運動制御
の分子機構などについてはまだなぞが多い。長い進化・淘汰の末に極度に最適化された運動器官であると考えられ、その自動制御機構を人工的な系で模倣するこ
とが私の研究の目的であるが、上の理由で、医生物学的な面からもしくみ解明が急務となっている。
対象とするもののサイズが小さいこと、これが研究上の一番の障害となっている。運動の観察は、生きた状態で行わなければならず、当然、光学顕微鏡などを
使わざるを得ないが、分解能の点で観察できる現象には限界がある。こういった手法上の障壁をまずは克服しつつ研究することになる。2つの例を紹介する。1
つは高い精度で運動を解析するシステムで、ウニ精子のベン毛に付着させたマイクロビーズの位置を数十pm(10-12メー
トル)もの精度で計測する装置を大学院生の野田直紀君が完成させた(図S- 3)(6)。 これは水分子の半分ほどのスケールで恐ろしく高い精度である。分子スケー
ルどころか、サブ原子スケールの分子ゆらぎや振動を計測できる世界最高水準の計測精度である。ダイニンが振幅8nmの規則正しい振動をする特殊なモーター
分子であることはわかっていたが、その周期にゆるやかなゆらぎがあることもわかってきた。もう1つの例は、顕微鏡下での運動開始実験である。ウニ精子は通
常は40Hzほどで運動する(図S-4)が、この運動が
ATP加水分解反応(エネルギー供給源)とどのように関係しているのかを調べた(図S-5)。
これ
は谷知己氏(現・北大准教授)が考案した装置、名付けるならば『紫外線照射兼高速画像取り込み装置』を使ったもので、紫外線照射で分解してATPを作る
caged-ATPという化合物を使った実験である(7)。ATPが供給されて1/50秒ほど遅れて運動が開始する
こと、止まっていた運動は助走することもな
く、または、リセットされるのでもなく、何とそのままの姿で突然始まることがわかった。この観察からダイニン・ADPという反応中間体が運動に関わってい
ること、ベン毛のくねり運動は屈曲の曲り角度で刻々とフィードバック制御されていることなどがわかった。こういった光学顕微鏡を使った解析手法は、まだま
だ無限の可能性を秘めているのではないかと思う。私たちの研究室では、解析の目的に応じて、こういった光学顕微鏡技術と最新光エレクトロニクス手法の組み
合わせを模索することが、最初の重要な仕事となっている。さらに、この数年程前から、光学顕微鏡では探れない微細構造の変化にも着目し、AFM(8)やX線回折
の技術も使い始めている。X線解析の手法は、タンパク結晶や筋繊維など整然とした構造を持つものでその能力を大いに発揮する。ベン毛や繊毛を何万本と集め
て一束にし、それらを整然と並べることがまずは必須であるが、なかなかその解決方法が見つからなかった。昨年、やっとこの課題を流動配向という新技術で解
決し、SPring-8や高エネ研の専門家との共同研究によって、ベン毛の構造解析が可能となった(図3、図S-6)。チューブリンやダイニンなどのタン
パク質がベン毛の中でどのように配置し、それがどのような構造変化をするのか、世界ではじめて解析できるようになった。ATPでダイニンの配置が平行型か
らラセン型へとダイナミックに変わるという証拠(図S-7)も得られるようになって来た。この構造変化が運動制御のしくみに深く関わっているのではないか
と期待している。
研究の対象が小さくて扱いにくい。その分、従来型の手法に何かひと工夫するか、あるいは、全くこれまでにない新しい手法を創出しなければ、次の段階には
進めないことが多い。外見は大変そうな分野かも知れないが、新しい手法は確実に何らかの新しい知見をもたらし、それが思いもよらぬ方向へと研究を展開させ
ることが多いのも事実である。これは、よくよく考えると特別なことではない。どの研究分野にも共通して変わらない事実であ
注)文中の '図S-' で始まる図番号は、下のリンクサイトへアクセスすると表示できます。動画なども用意してありますので、お楽しみください。http://www.bio.chuo-u.ac.jp/nano/G2007/
図 2.9+ 2構造。多くの運動性のベン毛・繊毛に共通する構造。

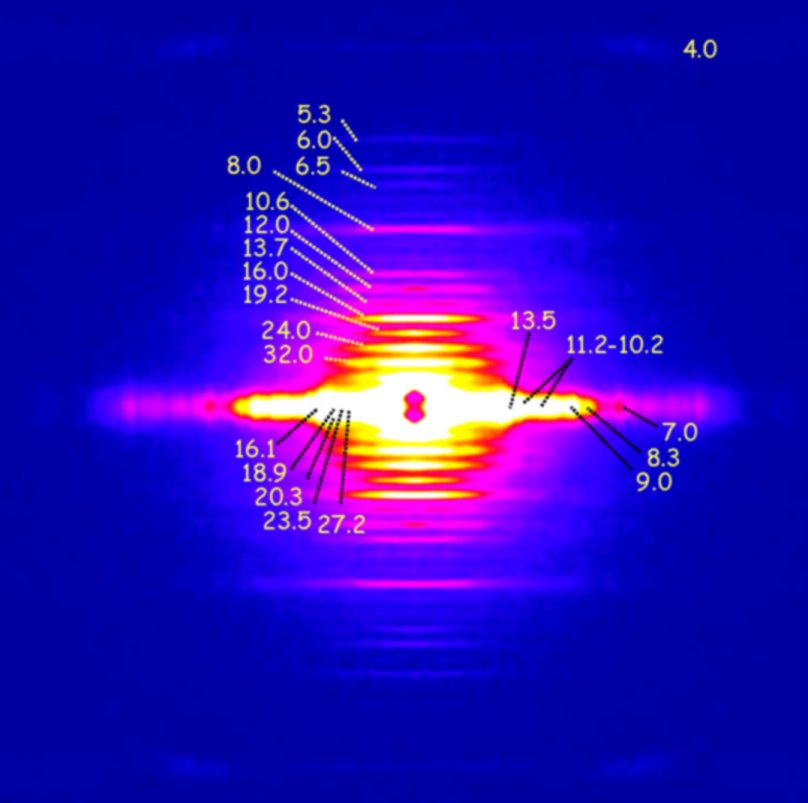
図3. ウニ精子ベン毛のX線回折。4nmのチューブリン分子の構造をはじめ、多くの周期性構造が見られる。数字は、周期構造の周期を示す。上下 が、ベン毛の長軸方向の情報。
1. Baldauf (2003) The deep roots of eukaryotes. Science, 300, 1703-1706.
2. Mitchell (2004) Speculations on the evolution of 9+2 organells and the role of central pair microtubules. Biol. Cell, 96, 691-696.
3. Banado, Mitsu, Beales & Katsanis (2006) The ciliopathies: An emerging class of human genetic disorders. Annu. Rev. Genomics Hum, Genet, 7, 125-148.
5. Fliegauf, Benzing & Omram (2007) When cilia go bad: cilia defects and ciliopathies. Nature Rev. Mole. Cell Biol., 8, 880-893.
6. Noda & Kamimura (2008) A new microscope optics for laser darkfield illumination applied to high precision two dimensional measurement of specimen displacement. Rev. Sci. Inst. (in press).
7. Tani & Kamimura (1998) Reactivation of sea-urchin sperm flagella induced by rapid photolysis of caged ATP. J. exp. Biol., 201, 1493-1503.
8. Sakaibara, Kunioka, Yamada & Kamimura (2004) Diameter oscillation of axonemes in sea-uechin sperm flagella. Biophys. J., 86, 346-352.